Antimicrobial Peptides in Orthopaedics: Current Status and Future Prospects
Meghan McCaskey, BA; Zbigniew Gugala, MD, PhD
Department of Orthopaedic Surgery & Rehabilitation, University of Texas Medical Branch; Galveston, TX, USA
Corresponding Author:Zbigniew Gugala, MD, PhD, Department or Orthopaedic Surgery & Rehabilitation, University of Texas Medical Branch, 301 University Blvd, Galveston, TX 77555; USA; zgugala@utmb.edu
DOI: 10.18600/toj.020113
INTRODUCTION
With the rise of antibiotic-resistant bacteria and an increased incidence of periprosthetic joint infection (PJI) [1], a need has arisen for novel approaches to fighting infections, in particular osteomyelitis. This need is accentuated by the emergence of multidrug-resistant bacteria resulting from the overuse of antibiotics [2]. The need becomes increasingly important as 0.2-5.0% of cases develop PJI where the most common cause of infection is Staphylococcus strains [3]. In particular, multiresistant Staphylococcus aureus and vancomycin-resistant Enterococcus faecium pose major risks for infection in terms of osteomyelitis, as they are recognized to cause infections in bones and joints [1,2,4]. One of the main concerns surrounding these multidrug-resistant bacteria is the ability to form biofilms on the surface of orthopaedic implants. Biofilms are a highly structured community of bacteria that are from 10 to 1,000 times more resistant to drug therapies than planktonic bacteria [3,5,6]. Because of the resistant nature of these bacteria and the capability of biofilm formation, treatment and control of the infection are particularly challenging [1]. One appealing approach to combating these multiresistant bacteria is through the use of antimicrobial peptides. Antimicrobial peptides are naturally expressed and utilized by the innate immune system as an additional layer of defense against pathogens [7]. Antimicrobial peptides are usually cationic and are therefore selective towards particular cell membrane compositions. These peptides additionally serve as local agents and prove to have very little systemic impact, making them excellent candidates for therapy [5]. There is great diversity between the structure and mechanisms of action of these antimicrobial peptides, which can impact the spectrum of microbes against which they are active.
Cathelicidins and defensins are 2 classes of antimicrobial peptides expressed in humans that exhibit therapeutic promise. Cathelicidin peptides exhibit a great deal of diversity in both their sequence and structure, but still are active as immunomodulators and/or antimicrobial agents [8]. They are cationic and small (12-97 residue) peptides that possess anti-infective properties [8]. One well studied cathelicidin is LL-37, which is expressed in humans as part of the innate immune response. Expression of LL-37 is induced by infection; utilizing this knowledge provides interesting therapeutic or diagnostic opportunities [5,8].
Unlike cathelicidins, defensins present a conserved scaffold of an antiparallel β-sheet while still maintaining high diversity in the sequence [9]. Although α-defensins show promise as a diagnostic tool [10,11]], the focus for therapeutic application lies on β-defensins because they have a naturally wider tissue distribution, constitutively expressed or induced by bacteria or proinflammatory cytokines, and are able to act as immune effectors [9]. Beta defensins are known to be produced by osteocytes and osteoblasts—an attribute which adds to their appeal as a therapeutic option for osteomyelitis and PJI [5]. The expression of β-defensins is induced by infection in the osteoblasts and osteocytes [12]. These peptides are active at low concentrations (10-100 μg/mL) and together are active against Gram-positive, Gram-negative bacteria, mycobacteria, fungi, and enveloped viruses [13]. Beta defensins contain 38-42 residues, including 6 cysteine residues linking C1 to C5, C2 to C4, and C3 to C6 [8]. The remainder of the sequence is highly variable, demonstrating the conservation of the scaffold with sequence diversity. Beta-defensins are known to represent an important part in the innate immune system in humans. Immunological activity can be observed in cases of osteoradionecrosis (ORN), where viable bone close to the necrotic bone expresses β-defensin-1 (HBD-1), HBD-2, and HBD-3 to provide immunological functions [7]. These peptides because of their residence in bone tissue, in addition to having broad antimicrobial activity, chemical resistance, and lack of antigenicity, provide further evidence of being useful therapeutic agents in terms of osteomyelitis [14].
Current Status
Diagnosis of osteomyelitis is often difficult and unclear regardless of routine inflammatory laboratory values, radiologic studies, and aspirate fluid and culture analysis [15]. There is evidence that both α-defensins and LL-37 could be valuable in the diagnostic process. In a recent study, synovial fluid levels of α-defensins demonstrate nearly 100% sensitivity and specificity for diagnosis of PJI [15]. Virulence did not impact the magnitude of response by α-defensins, thereby eliminating concerns surrounding low virulence and false negatives [16]. Alpha-defensins gain additional credibility in diagnostic testing for osteomyelitis and PJI by eliminating virulence as a diagnostic hurdle. It is important to note that the study included patients with antibiotic administration and/or systemic inflammatory arthropathies, which both frequently frustrate the diagnostic process [15] LL-37 and HBD-3 appear to be equally impressive candidates in terms of diagnostic capabilities, both demonstrating significant local upregulation in patients with PJI [5].
In addition to being able to use these peptides as diagnostic tools, HBD-2 and, more recently, HBD-3 demonstrates the ability to be biocompatible and potentially useful as therapeutic agents [17]. Attachment of HBD-2 to implants as an antimicrobial technique has shown to prevent infection with 100% killing of E. coli (used as a Gram-negative bacteria test) observed up to 2 hours post-introduction of implant to bacteria, and 60% killing observed 6 hours post-introduction [2]. Although HBD-2 is active against Gram-negative bacteria, it is not active against Gram-positive bacteria or fungi, such as yeast [4]. HBD-2’s inactivity against Gram-positive bacteria presents a problem, with 80% of osteomyelitis cases stemming from Staphylococcus aureus, a Gram-positive bacterial strain [18]. HBD-2 also does not retain antimicrobial functionality at physiological salt concentrations, which discourages its use as a therapeutic agent [19]. Demonstrating the concept that defensins can be utilized to coat implants and provide antimicrobial effects is an integral step toward developing novel techniques for combating infection. Unlike HBD-2, HBD-3 is known to work over a broad spectrum of organisms. HBD-3 is the most cationic β-defensin, as well as being the most potent antibacterial agent displaying activity against gram-negative bacteria, gram-positive bacteria, fungi, such as yeast or Saccharomyces cerevisiae, and bacterial complexes, such as Burkholderia cepacea [4,20,21]. The activity against Staphylococcus aureus and against multiple other multiresistant bacterial strains is the most important antimicrobial activity exhibited by HBD-3 [18]. HBD-3 is also known to have greater stability relative to HBD-2 and is able to maintain its antimicrobial function at physiological salt conditions, making it a promising candidate for therapeutic use [22]. In addition to stability and salt-insensitivities, HBD-3 displays no hemolytic activity using concentrations up to 500 μg/mL [12]. This absence of hemolytic activity indicates that HBD-3 could be a safe therapeutic option as it is not active against eukaryotic cells, more specifically, erythrocytes. Higher concentrations of HBD-3 yield more potent effects on the multiresistant bacteria, which potentially could be utilized without harming the patient.
Finally, HBD-3 demonstrates the capability to inhibit biofilm formation [3,6,23]. Because biofilm formation is a dynamic process, it is important to look at which stage of formation HBD-3 inhibits. By looking at dltB and icaA levels, 2 bacterial genes known to assist in biofilm formation, some mechanistic details can be revealed [23]. HBD-3 appears to stimulate icaA upregulation but appears to not have a significant impact on dltB. This effect is significant as vancomycin and clindamycin, 2 commonly used antibiotics, illicit a moderate increase in icaA transcription but significantly induce the upregulation of dltB transcription [23]. The differences in regulation patterns of icaA and dltB indicate that the mechanism of inhibition demonstrates a more complicated effect of HBD-3 compared to vancomycin and clindamycin on icaA and dltB regulation [23]. Even though the regulation patterns of vancomycin, clindamycin, and HBD-3 are different, the effectiveness against biofilms does not appear to be impacted. As the concentration of vancomycin, clindamycin, and HBD-3 increases, the bacterial colonies decrease during the biofilm adhesion phase, or first phase, of biofilm formation [6]. In the biofilm formation phase, when the concentration of HBD-3 and clindamycin are greater than the MIC, there is significant inhibition of biofilm formation. This finding illustrates more sensitivity than vancomycin, where the concentration necessary to inhibit biofilm formation increases to between 2 times and 4 times the MIC [6]. In the biofilm initial mature phase, inhibition potency starts to waver between the 3 antibacterials, where HBD-3 significantly reduces the biofilm area in both the 2 times and 4 times the MIC groups. Clindamycin also reduced the biofilm area in both the 2 times and 4 times the MIC groups; however it exhibited less potency on already formed biofilms than HBD-3 [6]. Vancomycin did not affect the biofilm area, exhibiting even less potency on already formed biofilms than both clindamycin and HBD-3. Because of the stability of HBD-3, the broad-spectrum action displayed by HBD-3, and the impact HBD-3 has on biofilm formation, HBD-3 is considered to be the most promising class of defensin antimicrobial peptides, especially in terms of therapy for orthopaedic implant-related infections [3,6].
With the rise of antibiotic-resistant bacteria and an increased incidence of periprosthetic joint infection (PJI) [1], a need has arisen for novel approaches to fighting infections, in particular osteomyelitis. This need is accentuated by the emergence of multidrug-resistant bacteria resulting from the overuse of antibiotics [2]. The need becomes increasingly important as 0.2-5.0% of cases develop PJI where the most common cause of infection is Staphylococcus strains [3]. In particular, multiresistant Staphylococcus aureus and vancomycin-resistant Enterococcus faecium pose major risks for infection in terms of osteomyelitis, as they are recognized to cause infections in bones and joints [1,2,4]. One of the main concerns surrounding these multidrug-resistant bacteria is the ability to form biofilms on the surface of orthopaedic implants. Biofilms are a highly structured community of bacteria that are from 10 to 1,000 times more resistant to drug therapies than planktonic bacteria [3,5,6]. Because of the resistant nature of these bacteria and the capability of biofilm formation, treatment and control of the infection are particularly challenging [1]. One appealing approach to combating these multiresistant bacteria is through the use of antimicrobial peptides. Antimicrobial peptides are naturally expressed and utilized by the innate immune system as an additional layer of defense against pathogens [7]. Antimicrobial peptides are usually cationic and are therefore selective towards particular cell membrane compositions. These peptides additionally serve as local agents and prove to have very little systemic impact, making them excellent candidates for therapy [5]. There is great diversity between the structure and mechanisms of action of these antimicrobial peptides, which can impact the spectrum of microbes against which they are active.
Cathelicidins and defensins are 2 classes of antimicrobial peptides expressed in humans that exhibit therapeutic promise. Cathelicidin peptides exhibit a great deal of diversity in both their sequence and structure, but still are active as immunomodulators and/or antimicrobial agents [8]. They are cationic and small (12-97 residue) peptides that possess anti-infective properties [8]. One well studied cathelicidin is LL-37, which is expressed in humans as part of the innate immune response. Expression of LL-37 is induced by infection; utilizing this knowledge provides interesting therapeutic or diagnostic opportunities [5,8].
Unlike cathelicidins, defensins present a conserved scaffold of an antiparallel β-sheet while still maintaining high diversity in the sequence [9]. Although α-defensins show promise as a diagnostic tool [10,11]], the focus for therapeutic application lies on β-defensins because they have a naturally wider tissue distribution, constitutively expressed or induced by bacteria or proinflammatory cytokines, and are able to act as immune effectors [9]. Beta defensins are known to be produced by osteocytes and osteoblasts—an attribute which adds to their appeal as a therapeutic option for osteomyelitis and PJI [5]. The expression of β-defensins is induced by infection in the osteoblasts and osteocytes [12]. These peptides are active at low concentrations (10-100 μg/mL) and together are active against Gram-positive, Gram-negative bacteria, mycobacteria, fungi, and enveloped viruses [13]. Beta defensins contain 38-42 residues, including 6 cysteine residues linking C1 to C5, C2 to C4, and C3 to C6 [8]. The remainder of the sequence is highly variable, demonstrating the conservation of the scaffold with sequence diversity. Beta-defensins are known to represent an important part in the innate immune system in humans. Immunological activity can be observed in cases of osteoradionecrosis (ORN), where viable bone close to the necrotic bone expresses β-defensin-1 (HBD-1), HBD-2, and HBD-3 to provide immunological functions [7]. These peptides because of their residence in bone tissue, in addition to having broad antimicrobial activity, chemical resistance, and lack of antigenicity, provide further evidence of being useful therapeutic agents in terms of osteomyelitis [14].
Current Status
Diagnosis of osteomyelitis is often difficult and unclear regardless of routine inflammatory laboratory values, radiologic studies, and aspirate fluid and culture analysis [15]. There is evidence that both α-defensins and LL-37 could be valuable in the diagnostic process. In a recent study, synovial fluid levels of α-defensins demonstrate nearly 100% sensitivity and specificity for diagnosis of PJI [15]. Virulence did not impact the magnitude of response by α-defensins, thereby eliminating concerns surrounding low virulence and false negatives [16]. Alpha-defensins gain additional credibility in diagnostic testing for osteomyelitis and PJI by eliminating virulence as a diagnostic hurdle. It is important to note that the study included patients with antibiotic administration and/or systemic inflammatory arthropathies, which both frequently frustrate the diagnostic process [15] LL-37 and HBD-3 appear to be equally impressive candidates in terms of diagnostic capabilities, both demonstrating significant local upregulation in patients with PJI [5].
In addition to being able to use these peptides as diagnostic tools, HBD-2 and, more recently, HBD-3 demonstrates the ability to be biocompatible and potentially useful as therapeutic agents [17]. Attachment of HBD-2 to implants as an antimicrobial technique has shown to prevent infection with 100% killing of E. coli (used as a Gram-negative bacteria test) observed up to 2 hours post-introduction of implant to bacteria, and 60% killing observed 6 hours post-introduction [2]. Although HBD-2 is active against Gram-negative bacteria, it is not active against Gram-positive bacteria or fungi, such as yeast [4]. HBD-2’s inactivity against Gram-positive bacteria presents a problem, with 80% of osteomyelitis cases stemming from Staphylococcus aureus, a Gram-positive bacterial strain [18]. HBD-2 also does not retain antimicrobial functionality at physiological salt concentrations, which discourages its use as a therapeutic agent [19]. Demonstrating the concept that defensins can be utilized to coat implants and provide antimicrobial effects is an integral step toward developing novel techniques for combating infection. Unlike HBD-2, HBD-3 is known to work over a broad spectrum of organisms. HBD-3 is the most cationic β-defensin, as well as being the most potent antibacterial agent displaying activity against gram-negative bacteria, gram-positive bacteria, fungi, such as yeast or Saccharomyces cerevisiae, and bacterial complexes, such as Burkholderia cepacea [4,20,21]. The activity against Staphylococcus aureus and against multiple other multiresistant bacterial strains is the most important antimicrobial activity exhibited by HBD-3 [18]. HBD-3 is also known to have greater stability relative to HBD-2 and is able to maintain its antimicrobial function at physiological salt conditions, making it a promising candidate for therapeutic use [22]. In addition to stability and salt-insensitivities, HBD-3 displays no hemolytic activity using concentrations up to 500 μg/mL [12]. This absence of hemolytic activity indicates that HBD-3 could be a safe therapeutic option as it is not active against eukaryotic cells, more specifically, erythrocytes. Higher concentrations of HBD-3 yield more potent effects on the multiresistant bacteria, which potentially could be utilized without harming the patient.
Finally, HBD-3 demonstrates the capability to inhibit biofilm formation [3,6,23]. Because biofilm formation is a dynamic process, it is important to look at which stage of formation HBD-3 inhibits. By looking at dltB and icaA levels, 2 bacterial genes known to assist in biofilm formation, some mechanistic details can be revealed [23]. HBD-3 appears to stimulate icaA upregulation but appears to not have a significant impact on dltB. This effect is significant as vancomycin and clindamycin, 2 commonly used antibiotics, illicit a moderate increase in icaA transcription but significantly induce the upregulation of dltB transcription [23]. The differences in regulation patterns of icaA and dltB indicate that the mechanism of inhibition demonstrates a more complicated effect of HBD-3 compared to vancomycin and clindamycin on icaA and dltB regulation [23]. Even though the regulation patterns of vancomycin, clindamycin, and HBD-3 are different, the effectiveness against biofilms does not appear to be impacted. As the concentration of vancomycin, clindamycin, and HBD-3 increases, the bacterial colonies decrease during the biofilm adhesion phase, or first phase, of biofilm formation [6]. In the biofilm formation phase, when the concentration of HBD-3 and clindamycin are greater than the MIC, there is significant inhibition of biofilm formation. This finding illustrates more sensitivity than vancomycin, where the concentration necessary to inhibit biofilm formation increases to between 2 times and 4 times the MIC [6]. In the biofilm initial mature phase, inhibition potency starts to waver between the 3 antibacterials, where HBD-3 significantly reduces the biofilm area in both the 2 times and 4 times the MIC groups. Clindamycin also reduced the biofilm area in both the 2 times and 4 times the MIC groups; however it exhibited less potency on already formed biofilms than HBD-3 [6]. Vancomycin did not affect the biofilm area, exhibiting even less potency on already formed biofilms than both clindamycin and HBD-3. Because of the stability of HBD-3, the broad-spectrum action displayed by HBD-3, and the impact HBD-3 has on biofilm formation, HBD-3 is considered to be the most promising class of defensin antimicrobial peptides, especially in terms of therapy for orthopaedic implant-related infections [3,6].


Future Prospects
HBD-3 stands out amongst other peptides as a good candidate for combating orthopaedic implant-related infections and osteomyelitis. The amino acid sequence along with the secondary structure created by the particular amino acids is illustrated by Figure 1 [22]. A 3-dimensional model of HDB-3 is depicted in Figure 2, illustrating the secondary structure present throughout HBD-3 as well as indicating the positions of the 3 disulfide bonds indicated in yellow [22]. It is apparent that there is no tertiary structure present in HBD-3, which introduces options in terms of making the peptide itself. There are 2 main ways to obtain HBD-3, peptide synthesis or vector recombination. As mentioned above, the lack of tertiary structure and the small size of HBD-3 make peptide synthesis a strong possibility in terms of obtaining viable peptide. Recombination techniques can also be utilized for creating HBD-3, although they are more complicated because the short peptides are on the limit of having adequate expression, as well as the peptide itself being toxic to the bacteria cells [20]. Even though it is markedly more complicated, there are ways to accomplish expression of HBD-3 in bacteria as charted below in Table 1 [20]. The chart shows the optimal conditions for successful recombination, including bacteria type, fusion partner protein, method of fusion cleavage, and expected expression level. With this information HBD-3 can be produced. Once the production and purification of HBD-3 is complete, testing to determine antimicrobial activity can be conducted by introducing HBD-3 to a variety of bacterial strains and observing the viability of the bacteria.
HBD-3 stands out amongst other peptides as a good candidate for combating orthopaedic implant-related infections and osteomyelitis. The amino acid sequence along with the secondary structure created by the particular amino acids is illustrated by Figure 1 [22]. A 3-dimensional model of HDB-3 is depicted in Figure 2, illustrating the secondary structure present throughout HBD-3 as well as indicating the positions of the 3 disulfide bonds indicated in yellow [22]. It is apparent that there is no tertiary structure present in HBD-3, which introduces options in terms of making the peptide itself. There are 2 main ways to obtain HBD-3, peptide synthesis or vector recombination. As mentioned above, the lack of tertiary structure and the small size of HBD-3 make peptide synthesis a strong possibility in terms of obtaining viable peptide. Recombination techniques can also be utilized for creating HBD-3, although they are more complicated because the short peptides are on the limit of having adequate expression, as well as the peptide itself being toxic to the bacteria cells [20]. Even though it is markedly more complicated, there are ways to accomplish expression of HBD-3 in bacteria as charted below in Table 1 [20]. The chart shows the optimal conditions for successful recombination, including bacteria type, fusion partner protein, method of fusion cleavage, and expected expression level. With this information HBD-3 can be produced. Once the production and purification of HBD-3 is complete, testing to determine antimicrobial activity can be conducted by introducing HBD-3 to a variety of bacterial strains and observing the viability of the bacteria.
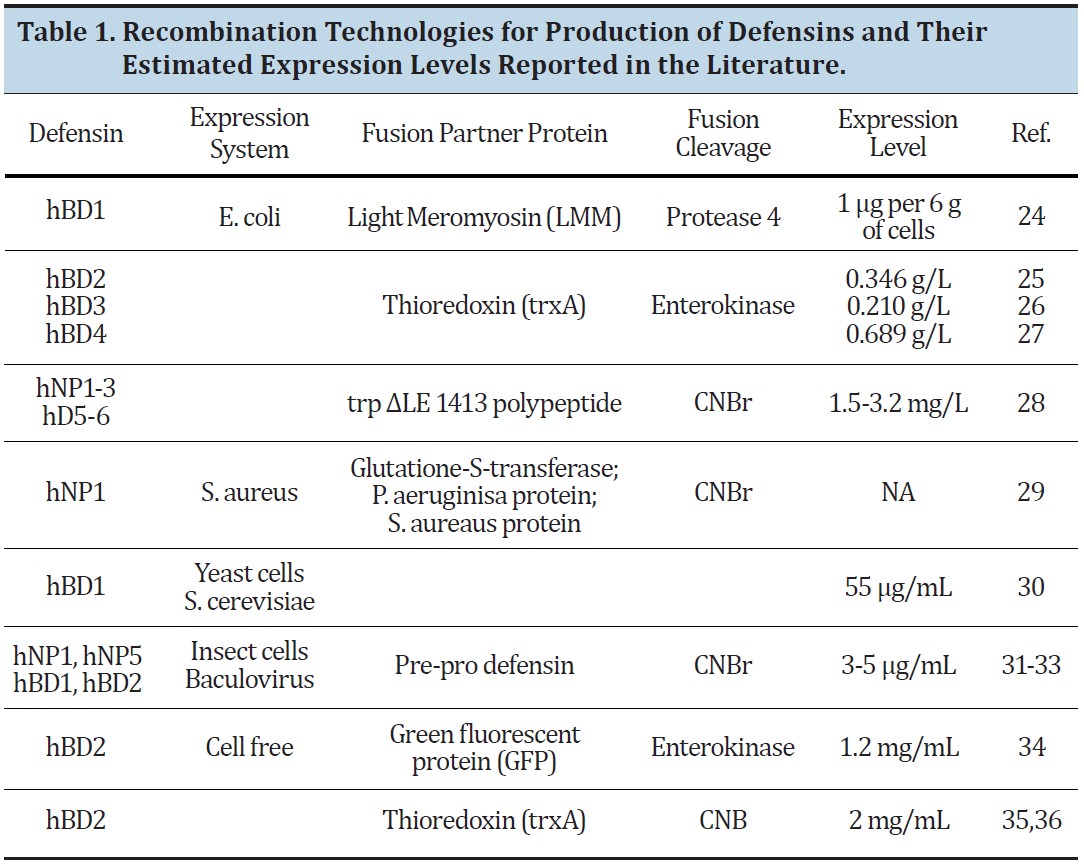
After the antimicrobial activity of the produced HBD-3 is confirmed, coating orthopaedic implants with bioactive peptides can be tested [2,17]. The use of HBD-3 as an antimicrobial coating on orthopaedic implants to combat biofilm formation of multidrug-resistant bacteria in the body can then be studied. The potential exists to further lower the incidence of osteomyelitis and implant-related infections without further inducing bacterial resistance. HBD-3 shows promise in as a bioactive agent that can prevent bacterial colonization and biofilm formation on orthopaedic implants [2].
REFERENCES
[1] Cunningham R, Cockayne A, Humphreys H. Clinical and molecular aspects of the pathogenesis of Staphylococcus aureus bone and joint infections. J Med Microbiol. 1996;44:157-64.
[2] Pfeufer N, Hofmann-Peiker K, Mühle M, Warnke P, Weigel M, Kleine M. Bioactive coating of titanium surfaces with recombinant human β-defensin-2 (rHuβD2) may prevent bacterial colonization in orthopaedic surgery. J Bone Joint Surg. 2011;93:840-6.
[3] Zhu C, Tan H, Cheng T, Shen H, Shao J, Guo Y, Shi S, Zhang X. Human β-defensin 3 inhibits antibiotic-resistant Staphylococcus biofilm formation. J Surg Res. 2013;183:204-13.
[4] Schneider J, Unholzer A, Schalter M, Schäfer-Korting M, Korting H. Human defensins. J Mol Med. 2005;83:587-95.
[5] Gollwitzer H, Dombrowski Y, Prodinger P, Peric M, Summer B, Hapfelmeier A, Dipl-Stat, Saldamli B, Pankow F, Eisenhart-Rothe R, Imhoff A, Schauber J, Thomas P, Burgkart R, Banke I. Antimicrobial peptides and proinflammatory cytokines in periprosthetic joint infection. J Bone Joint Surg. 2013;95:644-51.
[6] Huang Q, Yu H, Liu G, Huang X, Zhang L, Zhou Y, Chen J, Lin F, Wang Y, Fei J. Comparison of the Effects of Human β-defensin 3, Vancomycin, and clindamycin on Staphylococcus aureus biofilm formation. Orthopedics. 2012;35:e53-e60.
[7] Warnke P, Springer I, Russo P, Wiltfang J, Essig H, Kosmahl M, Sherry E, Acil Y. Innate immunity in human bone. Bone. 2006;38:400-8.
[8] Mookherjee N, Brown K, Hancock R. Cathelicidins. Handbook of Biologically Active Peptides: Chapter 15. 2013;77-84.
[9] Antcheva N, Guida F, Tossi A. Defensins. Handbook of biologically active peptides: Chapter 18. 2013;101-18.
[10] Deirmengian C, Kardos K, Kilmartin P, Cameron A, Schiller K, Booth RE Jr, Parvizi J. The alpha-defensin test for periprosthetic joint infection outperforms the leukocyte esterase test strip. Clin Orthop Relat Res. 2015;473(1):198-203.
[11] Bonanzinga T, Zahar A, Dütsch M, Lausmann C, Kendoff D, Gehrke T. How reliable is the alpha-defensin immunoassay test for diagnosing periprosthetic joint Infection? A prospective study. Clin Orthop Relat Res. 2017;475(2):408-15.
[12] Harder J, Bartels J, Christophers E, Schröder J. Isolation and characterization of human β-defensin-3, a novel human inducible peptide antibiotic. J Biol Chem. 2001;276:5707-13.
[13] Ganz T, Lehrer R. Defensins. Current Biology. 1994;6:584-9.
[14] Ganz T, Lehrer R. Defensins. Pharmac Ther. 1995;66:191-205. [15] Potter B. Alpha-defensin—the biggest thing in joint replacement infections since prophylactic antibiotics. Clin Orthop Relat Res. 2015;473:3105-7.
[16] Springer B. The alpha-defensin test for periprosthetic joint infection responds to a wide spectrum of organisms. Clin Orthop Relat Res. 2015;473:2236-7.
[17] Warnke P, Voss E, Russo P, Stephens S, Kleine M, Terheyden H, Liu Q. Antimicrobial peptide coating of dental implants: biocompatibility assessment of recombinant human beta defensin-2 for human cells. Int J Oral Maxillof Implants. 2013;28:982-8.
[18] Varoga D, Wruck C, Tohidnezhad M, Brandenburg L, Paulsen F, Mentlein R, Seekamp A, Besch L, Pufe T. Osteoblasts participate in the innate immunity of the bone by producing human beta defensin-3. Histochem Cell Biol. 2009;131:207-18.
[19] Bals R, Wang X, Wu Z, Freeman T, Bafna V, Zasloff M, Wilson J. Human β-defensin 2 is a salt-sensitive peptide antibiotic expressed in human lung. J Clin Invest. 1998;102:874-80.
[20] Pazgier M, Li X, Lu W, Lubkowski J. Human Defensins: Synthesis and structural properties. Curr Pharml Des. 2007;13:3096-118.
[21] Varoga D, Pufe T, Harder J, Meyer-Hoffert U, Mentlein R, Schröder J, Petersen W, Tillmann B, Proksch E, Goldring M, Paulsen F. Production of endogenous antibiotics in articular cartilage. Arthritis Rheum. 2004;50:3526-34.
[22] Dhople V, Krukemeyer A, Ramamoorthy A. The human beta-defensin-3, an antibacterial peptide with multiple biological functions. Biochim Biophys Acta. 2006;1758:1499-512.
[23] Huang Q, Fei J, Yu H, Giu Y, Huang X. Effects of human β-defensin-3 on biofilm formation-regulating genes dltB and icaA in Staphylococcus aureus. Mol Med Rep. 2014;10:825-31.
[24] Cipakova I, Hostinova E, Gasperik J, Velebny V. High-level expression and purification of a recombinant hBD-1 fused to LMM protein in Escherichia coli. Protein Expr Purif. 2004;37:207-12.
[25] Xu Z, Zhong Z, Huang L, Peng L, Wang F, Cen P. High-level production of bioactive human beta-defensin-4 in Escherichia coli by soluble fusion expression. Appl Microbiol Biotechnol. 2006;72:471-9.
[26] Xu Z, Peng L, Zhong Z, Fang X, Cen P. High-level expression of a soluble functional antimicrobial peptide, human beta-defensin 2, in Escherichia coli. Biotechnol Prog. 2006;22:382-6.
[27] Huang L, Wang J, Zhong Z, Peng L, Chen H, Xu Z, et al. Production of bioactive human beta-defensin-3 in Escherichia coli by soluble fusion expression. Biotechnol Lett. 2006;28:627-32.
[28] Pazgier M, Lubkowski J. Expression and purification of recombinant human alpha-defensins in Escherichia coli. Protein Expr Purif. 2006;49(1):1-8.
[29] Piers KL, Brown MH, Hancock RE. Recombinant DNA procedures for producing small antimicrobial cationic peptides in bacteria. Gene. 1993;134:7-13.
[30] Cipakova I, Hostinova E. Production of the human-beta-defensin using Saccharomyces cerevisiae as a host. Protein Pept Lett. 2005;12:551-4.
[31] Valore EV, Park CH, Quayle AJ, Wiles KR, McCray PB Jr, Ganz T. Human beta-defensin-1: an antimicrobial peptide of urogenital tissues. J Clin Invest. 1998;101:1633-42.
[32] Valore EV, Martin E, Harwig SS, Ganz T. Intramolecular inhibition of human defensin HNP-1 by its propiece. J Clin Invest. 1996;97:1624-9.
[33] Porter EM, van Dam E, Valore EV, Ganz T. Broad-spectrum antimicrobial activity of human intestinal defensin 5. Infect Immun. 1997;65:2396-401.
[34] Xu Z, Chen H, Yin X, Xu N, Cen P. High-level expression of soluble human beta-defensin-2 fused with green fluorescent protein in Escherichia coli cell-free system. Appl Biochem Biotechnol. 2005;127:53-62.
[35] Chen H, Xu Z, Cen P. High-level expression of human betadefensin-2 gene with rare codons in E. coli cell-free system. Protein Pept Lett. 2006;13:155-62.
[36] Chen H, Xu Z, Xu N, Cen P. Efficient production of a soluble fusion protein containing human beta-defensin-2 in E. coli cell-free system. J Biotechnol. 2005;115:307-15.
[1] Cunningham R, Cockayne A, Humphreys H. Clinical and molecular aspects of the pathogenesis of Staphylococcus aureus bone and joint infections. J Med Microbiol. 1996;44:157-64.
[2] Pfeufer N, Hofmann-Peiker K, Mühle M, Warnke P, Weigel M, Kleine M. Bioactive coating of titanium surfaces with recombinant human β-defensin-2 (rHuβD2) may prevent bacterial colonization in orthopaedic surgery. J Bone Joint Surg. 2011;93:840-6.
[3] Zhu C, Tan H, Cheng T, Shen H, Shao J, Guo Y, Shi S, Zhang X. Human β-defensin 3 inhibits antibiotic-resistant Staphylococcus biofilm formation. J Surg Res. 2013;183:204-13.
[4] Schneider J, Unholzer A, Schalter M, Schäfer-Korting M, Korting H. Human defensins. J Mol Med. 2005;83:587-95.
[5] Gollwitzer H, Dombrowski Y, Prodinger P, Peric M, Summer B, Hapfelmeier A, Dipl-Stat, Saldamli B, Pankow F, Eisenhart-Rothe R, Imhoff A, Schauber J, Thomas P, Burgkart R, Banke I. Antimicrobial peptides and proinflammatory cytokines in periprosthetic joint infection. J Bone Joint Surg. 2013;95:644-51.
[6] Huang Q, Yu H, Liu G, Huang X, Zhang L, Zhou Y, Chen J, Lin F, Wang Y, Fei J. Comparison of the Effects of Human β-defensin 3, Vancomycin, and clindamycin on Staphylococcus aureus biofilm formation. Orthopedics. 2012;35:e53-e60.
[7] Warnke P, Springer I, Russo P, Wiltfang J, Essig H, Kosmahl M, Sherry E, Acil Y. Innate immunity in human bone. Bone. 2006;38:400-8.
[8] Mookherjee N, Brown K, Hancock R. Cathelicidins. Handbook of Biologically Active Peptides: Chapter 15. 2013;77-84.
[9] Antcheva N, Guida F, Tossi A. Defensins. Handbook of biologically active peptides: Chapter 18. 2013;101-18.
[10] Deirmengian C, Kardos K, Kilmartin P, Cameron A, Schiller K, Booth RE Jr, Parvizi J. The alpha-defensin test for periprosthetic joint infection outperforms the leukocyte esterase test strip. Clin Orthop Relat Res. 2015;473(1):198-203.
[11] Bonanzinga T, Zahar A, Dütsch M, Lausmann C, Kendoff D, Gehrke T. How reliable is the alpha-defensin immunoassay test for diagnosing periprosthetic joint Infection? A prospective study. Clin Orthop Relat Res. 2017;475(2):408-15.
[12] Harder J, Bartels J, Christophers E, Schröder J. Isolation and characterization of human β-defensin-3, a novel human inducible peptide antibiotic. J Biol Chem. 2001;276:5707-13.
[13] Ganz T, Lehrer R. Defensins. Current Biology. 1994;6:584-9.
[14] Ganz T, Lehrer R. Defensins. Pharmac Ther. 1995;66:191-205. [15] Potter B. Alpha-defensin—the biggest thing in joint replacement infections since prophylactic antibiotics. Clin Orthop Relat Res. 2015;473:3105-7.
[16] Springer B. The alpha-defensin test for periprosthetic joint infection responds to a wide spectrum of organisms. Clin Orthop Relat Res. 2015;473:2236-7.
[17] Warnke P, Voss E, Russo P, Stephens S, Kleine M, Terheyden H, Liu Q. Antimicrobial peptide coating of dental implants: biocompatibility assessment of recombinant human beta defensin-2 for human cells. Int J Oral Maxillof Implants. 2013;28:982-8.
[18] Varoga D, Wruck C, Tohidnezhad M, Brandenburg L, Paulsen F, Mentlein R, Seekamp A, Besch L, Pufe T. Osteoblasts participate in the innate immunity of the bone by producing human beta defensin-3. Histochem Cell Biol. 2009;131:207-18.
[19] Bals R, Wang X, Wu Z, Freeman T, Bafna V, Zasloff M, Wilson J. Human β-defensin 2 is a salt-sensitive peptide antibiotic expressed in human lung. J Clin Invest. 1998;102:874-80.
[20] Pazgier M, Li X, Lu W, Lubkowski J. Human Defensins: Synthesis and structural properties. Curr Pharml Des. 2007;13:3096-118.
[21] Varoga D, Pufe T, Harder J, Meyer-Hoffert U, Mentlein R, Schröder J, Petersen W, Tillmann B, Proksch E, Goldring M, Paulsen F. Production of endogenous antibiotics in articular cartilage. Arthritis Rheum. 2004;50:3526-34.
[22] Dhople V, Krukemeyer A, Ramamoorthy A. The human beta-defensin-3, an antibacterial peptide with multiple biological functions. Biochim Biophys Acta. 2006;1758:1499-512.
[23] Huang Q, Fei J, Yu H, Giu Y, Huang X. Effects of human β-defensin-3 on biofilm formation-regulating genes dltB and icaA in Staphylococcus aureus. Mol Med Rep. 2014;10:825-31.
[24] Cipakova I, Hostinova E, Gasperik J, Velebny V. High-level expression and purification of a recombinant hBD-1 fused to LMM protein in Escherichia coli. Protein Expr Purif. 2004;37:207-12.
[25] Xu Z, Zhong Z, Huang L, Peng L, Wang F, Cen P. High-level production of bioactive human beta-defensin-4 in Escherichia coli by soluble fusion expression. Appl Microbiol Biotechnol. 2006;72:471-9.
[26] Xu Z, Peng L, Zhong Z, Fang X, Cen P. High-level expression of a soluble functional antimicrobial peptide, human beta-defensin 2, in Escherichia coli. Biotechnol Prog. 2006;22:382-6.
[27] Huang L, Wang J, Zhong Z, Peng L, Chen H, Xu Z, et al. Production of bioactive human beta-defensin-3 in Escherichia coli by soluble fusion expression. Biotechnol Lett. 2006;28:627-32.
[28] Pazgier M, Lubkowski J. Expression and purification of recombinant human alpha-defensins in Escherichia coli. Protein Expr Purif. 2006;49(1):1-8.
[29] Piers KL, Brown MH, Hancock RE. Recombinant DNA procedures for producing small antimicrobial cationic peptides in bacteria. Gene. 1993;134:7-13.
[30] Cipakova I, Hostinova E. Production of the human-beta-defensin using Saccharomyces cerevisiae as a host. Protein Pept Lett. 2005;12:551-4.
[31] Valore EV, Park CH, Quayle AJ, Wiles KR, McCray PB Jr, Ganz T. Human beta-defensin-1: an antimicrobial peptide of urogenital tissues. J Clin Invest. 1998;101:1633-42.
[32] Valore EV, Martin E, Harwig SS, Ganz T. Intramolecular inhibition of human defensin HNP-1 by its propiece. J Clin Invest. 1996;97:1624-9.
[33] Porter EM, van Dam E, Valore EV, Ganz T. Broad-spectrum antimicrobial activity of human intestinal defensin 5. Infect Immun. 1997;65:2396-401.
[34] Xu Z, Chen H, Yin X, Xu N, Cen P. High-level expression of soluble human beta-defensin-2 fused with green fluorescent protein in Escherichia coli cell-free system. Appl Biochem Biotechnol. 2005;127:53-62.
[35] Chen H, Xu Z, Cen P. High-level expression of human betadefensin-2 gene with rare codons in E. coli cell-free system. Protein Pept Lett. 2006;13:155-62.
[36] Chen H, Xu Z, Xu N, Cen P. Efficient production of a soluble fusion protein containing human beta-defensin-2 in E. coli cell-free system. J Biotechnol. 2005;115:307-15.