Heterotopic Ossification: Breaking the Cellular and Molecular Barriers
Austin Dickerson, BA¹; Diana Hoang, BS¹; Eleanor L. Davis¹; Alan R. Davis, PhD¹·²·³; Elizabeth A. Olmsted-Davis, PhD¹·²·³
¹Center for Cell and Gene Therapy, Baylor College of Medicine, Texas Children’s Hospital, Houston Methodist Hospital; Houston, TX, USA ²Department of Pediatrics–Section Hematology/Oncology, Baylor College of Medicine; Houston, TX, USA ³Department of Orthopaedic Surgery, Baylor College of Medicine; Houston, TX, USA
Corresponding Author:Elizabeth Olmsted-Davis, PhD, Center for Cell and Gene Therapy, Baylor College of Medicine, One Baylor Plaza, N1010, Houston, TX 77030; USA; edavis@bcm.edu
DOI: 10.18600/toj.020112
INTRODUCTION
Heterotopic ossification (HO) is the de novo formation of bone in nonosseous tissues. This type of atypical growth can be a source of significant clinical symptoms, leading to pain, edema, and a decreased range of motion of the affected joint. HO can result from genetic diseases, such as fibrodyplasia ossificans progressiva (FOP), or from traumatic or neurogenic causes. Joint arthroplasty, trauma to extremities, burns, amputation, blast-related trauma, and central or peripheral nervous system injury are among the most commonly cited conditions resulting in HO [1-3]. HO comprises a disorder that is characterized by the transformation of progenitor cells to osteogenic cells and begins with a neuroinflammatory process that appears to recruit and/or expand chondrocyte, osteoblast, brown adipocyte, and glial cell progenitors [4,5]. The onset of HO occurs in a localized microenvironment within the soft tissues, and it includes a lowered oxygen tension, availability of micronutrients, and an ideal pH [6-8]. The growth of the new bone is preceded by the establishment of vascular networks mediated by angiogenic growth factors secreted from brown fat as well as the mobilization of brown fat, through induction of hypoxia-inducible factor 1 (HIF1) to create a normoxic rather than hypoxic microenvironment that directly interfaces with the hypoxic area [6,9,10]. The formation of bone is at the expense of the local soft tissue, which often starts to atrophy and eventually dies. Since the pathophysiology of HO is still relatively unknown, ongoing efforts to examine its manifestation and understand the underlying mechanisms are necessary to aid in assessing risk and preventing the onset of this disorder.
Clinical Manifestations of HO
HO Associated with Genetic Disorders
In rare circumstances, HO may have genetic etiology. One such condition is FOP. This rare disorder is characterized by a mutation of the ACVR1 gene, which encodes the bone morphogenetic protein (BMP) receptor 2/4 resulting in the extensive and progressive ossification of soft connective tissues [11]. The mutation is known to cause mild constitutive activation of the BMP signaling pathway, suggesting that BMPs play a crucial role in driving osteogenesis and HO. Its hereditary transmission is autosomal dominant with variable expression [11]. There is no known cure for FOP; however, studies are underway to further understand the developmental biology behind it to identify potential therapeutic targets and prevent the mutation from initiating extraskeletal bone formation.
HO and Upper Extremity Trauma
The upper extremity exhibits a number of regions that are especially susceptible to HO. The joints are the most commonly affected targets, with the elbow being a location particularly prevalent HO can also appear without direct injury to the bone. Evidence shows that trauma to the muscle itself can also induce HO [12]. Joint injuries, such as dislocations and fractures, can stimulate HO, as well as surgical procedures, such as arthroplasty. The joints of the upper extremity have relatively high incidence rates for HO compared to other types of injuries. Elbow fractures are estimated to occur at a rate of 28 per 100,000 persons in the United States, while dislocations occur at a rate of approximately 11 per 100,000 persons [13]. In the elbow, where HO can be the most debilitating, the specific type of injury sustained often influences the presence of heterotopic bone formation. Retrospective studies involving patients with surgically treated elbow fractures showed that floating elbow injury, fractures on both sides of the joint, had the highest prevalence of HO reaching 36% [13]. Distal humerus fractures and “terrible triad” injuries were also found to have significant levels of clinically relevant HO with incidence rates of 12% and 18%, respectively [13,14]. Distal humerus fractures and “terrible triad” injuries were also found to have significant levels of clinically relevant HO [14,15]. Severe complications can result from this atypical bone growth in the elbow, such as ulnar nerve compression and decreased range of motion. Three factors associated with the prediction of clinically significant HO are type of injury, time to surgery, and time to mobilization after surgery. Patients who were operated on sooner and were able to mobilize the elbow more quickly after surgery had the least significant levels of heterotopic bone growth [14,16]. Fractures accompanied by dislocation are also predictors of HO, as heterotopic growth often accompanies the healing of these injuries [14-16].
HO and Lower Extremity Trauma
Posttraumatic HO occurs in soft tissues often secondary to any type of musculoskeletal injury associated with blast injuries and burns. Posttraumatic HO is mostly associated with large joints of the body in the lower extremities, including hip and knee joints. It is reported that 55% of patients with hip fractures develop HO and that the incidence rate rises to 83% if open reduction and fixation of the fracture are performed [17]. Acetabular fractures and their surgical treatment cause the most concern, with up to 58% of cases involving HO [16]. Studies show that the choice of surgical approach affects the likelihood of heterotopic bone development [16]. In addition, HO still remains 1 of the most frequent complications following hip arthroplasty. According to various studies, HO is diagnosed in 5% to 90% of patients who have undergone total hip arthroplasty (THA) with varying degrees of ossification from small fragments of bone to complete ankyloses of the hip joint [18]. Among those THA patients, 11% suffer significant HO symptoms, such as pain and limited range of motion [18]. It is suggested that secondary changes in nerve physiology from a blast injury may be responsible for inducing HO.
HO Associated with Amputation and Burn
HO has been a documented complication of limb amputation. This condition is especially relevant for military combat injuries, where the rates of amputations are higher than the general population [19]. In recent United States military conflicts in Iraq and Afghanistan, the incidence rate of amputation is estimated at 5.29 per 100,000 deployed troops [20]. HO occurs after traumatic and combat-related amputations and has been frequently noted as a potential cause of residual limb pain in amputees [19]. HO has proven to be a problematic clinical complication especially for prosthetic wear. In 1 study, HO was present in as many as 63% of all residual limbs postamputation [19]. It was determined that amputation near the injury site as well as a blast mechanism of injury were both significant risk factors for HO development. HO occurring at the site of an amputation can lead to pain in the residual limb as well as other complications, such as poor fitting of the prosthesis. The most effective forms of combating amputation-related HO are prophylaxis with a nonsteroidal anti-inflammatory drug and surgical excision of the bone growth [19]. Less frequent is the appearance of HO in severe burn victims with a varying incidence rate of 0.2% to 5.6% [21]. The association is poorly understood. However, it is suggested that burn injuries involve the recruitment of inflammatory molecules which promote the osteogenic differentiation of local mesenchymal stem cells [21].
HO Associated with Neurogenic Disorders
Severe neurological damage, such as spinal cord injury (SCI), head injuries, strokes, and brain tumors, can result in HO. In SCI, the incidence rate of HO varies from 16% to 53% with all HO occurring below the level of the spinal cord lesion, most commonly at the hip [22]. HO also occurs in 10% to 20% of patients with a closed head injury for whom the ossification mostly develops on the affected side of the brain trauma or stroke [22,23]. Limb spasticity has also been seen to increase the risk of HO development; those patients with extensive neurogenic HO usually present with severe spasticity [22].
Cellular/Molecular Manifestations of HO
HO Progenitors in the Peripheral Nerves
A great deal of controversy has surrounded the actual nature of the osteoprogenitor for HO. It was originally assumed that the progenitor for HO would be the bone marrow mesenchymal stem cell, originally defined by the experiments of Pittenger et al. [24]. These authors showed that cells from bone marrow that preferentially adhered to plastic could be isolated and shown to differentiate to osteoblasts, chondrocytes, and adipocytes. However, early published reports indicated that the progenitors for HO were derived locally rather than from bone marrow [25,26]. Then 3 reports appeared, 1 describing the endothelial origin of osteoprogenitors for HO [27], another noting that the progenitor was similar to those of skeletal muscle [28], and the third noting the neural origin of HO osteoprogenitors [5]. Interestingly, although these studies disagreed on the tissue of origin, they agreed upon the characteristic markers exhibited by the osteoprogenitor, which included Tie2, SP7 (osterix), and PDGFRA. Despite this controversy, it is generally agreed that HO is preceded by a period of neuroinflammation [4,5], where substance P and TRPV1 are expressed. If either of these pathways are blocked (eg, substance P binding to its receptor or TRPV1), then heterotopic bone formation is substantially reduced [4,5].
Heterotopic ossification (HO) is the de novo formation of bone in nonosseous tissues. This type of atypical growth can be a source of significant clinical symptoms, leading to pain, edema, and a decreased range of motion of the affected joint. HO can result from genetic diseases, such as fibrodyplasia ossificans progressiva (FOP), or from traumatic or neurogenic causes. Joint arthroplasty, trauma to extremities, burns, amputation, blast-related trauma, and central or peripheral nervous system injury are among the most commonly cited conditions resulting in HO [1-3]. HO comprises a disorder that is characterized by the transformation of progenitor cells to osteogenic cells and begins with a neuroinflammatory process that appears to recruit and/or expand chondrocyte, osteoblast, brown adipocyte, and glial cell progenitors [4,5]. The onset of HO occurs in a localized microenvironment within the soft tissues, and it includes a lowered oxygen tension, availability of micronutrients, and an ideal pH [6-8]. The growth of the new bone is preceded by the establishment of vascular networks mediated by angiogenic growth factors secreted from brown fat as well as the mobilization of brown fat, through induction of hypoxia-inducible factor 1 (HIF1) to create a normoxic rather than hypoxic microenvironment that directly interfaces with the hypoxic area [6,9,10]. The formation of bone is at the expense of the local soft tissue, which often starts to atrophy and eventually dies. Since the pathophysiology of HO is still relatively unknown, ongoing efforts to examine its manifestation and understand the underlying mechanisms are necessary to aid in assessing risk and preventing the onset of this disorder.
Clinical Manifestations of HO
HO Associated with Genetic Disorders
In rare circumstances, HO may have genetic etiology. One such condition is FOP. This rare disorder is characterized by a mutation of the ACVR1 gene, which encodes the bone morphogenetic protein (BMP) receptor 2/4 resulting in the extensive and progressive ossification of soft connective tissues [11]. The mutation is known to cause mild constitutive activation of the BMP signaling pathway, suggesting that BMPs play a crucial role in driving osteogenesis and HO. Its hereditary transmission is autosomal dominant with variable expression [11]. There is no known cure for FOP; however, studies are underway to further understand the developmental biology behind it to identify potential therapeutic targets and prevent the mutation from initiating extraskeletal bone formation.
HO and Upper Extremity Trauma
The upper extremity exhibits a number of regions that are especially susceptible to HO. The joints are the most commonly affected targets, with the elbow being a location particularly prevalent HO can also appear without direct injury to the bone. Evidence shows that trauma to the muscle itself can also induce HO [12]. Joint injuries, such as dislocations and fractures, can stimulate HO, as well as surgical procedures, such as arthroplasty. The joints of the upper extremity have relatively high incidence rates for HO compared to other types of injuries. Elbow fractures are estimated to occur at a rate of 28 per 100,000 persons in the United States, while dislocations occur at a rate of approximately 11 per 100,000 persons [13]. In the elbow, where HO can be the most debilitating, the specific type of injury sustained often influences the presence of heterotopic bone formation. Retrospective studies involving patients with surgically treated elbow fractures showed that floating elbow injury, fractures on both sides of the joint, had the highest prevalence of HO reaching 36% [13]. Distal humerus fractures and “terrible triad” injuries were also found to have significant levels of clinically relevant HO with incidence rates of 12% and 18%, respectively [13,14]. Distal humerus fractures and “terrible triad” injuries were also found to have significant levels of clinically relevant HO [14,15]. Severe complications can result from this atypical bone growth in the elbow, such as ulnar nerve compression and decreased range of motion. Three factors associated with the prediction of clinically significant HO are type of injury, time to surgery, and time to mobilization after surgery. Patients who were operated on sooner and were able to mobilize the elbow more quickly after surgery had the least significant levels of heterotopic bone growth [14,16]. Fractures accompanied by dislocation are also predictors of HO, as heterotopic growth often accompanies the healing of these injuries [14-16].
HO and Lower Extremity Trauma
Posttraumatic HO occurs in soft tissues often secondary to any type of musculoskeletal injury associated with blast injuries and burns. Posttraumatic HO is mostly associated with large joints of the body in the lower extremities, including hip and knee joints. It is reported that 55% of patients with hip fractures develop HO and that the incidence rate rises to 83% if open reduction and fixation of the fracture are performed [17]. Acetabular fractures and their surgical treatment cause the most concern, with up to 58% of cases involving HO [16]. Studies show that the choice of surgical approach affects the likelihood of heterotopic bone development [16]. In addition, HO still remains 1 of the most frequent complications following hip arthroplasty. According to various studies, HO is diagnosed in 5% to 90% of patients who have undergone total hip arthroplasty (THA) with varying degrees of ossification from small fragments of bone to complete ankyloses of the hip joint [18]. Among those THA patients, 11% suffer significant HO symptoms, such as pain and limited range of motion [18]. It is suggested that secondary changes in nerve physiology from a blast injury may be responsible for inducing HO.
HO Associated with Amputation and Burn
HO has been a documented complication of limb amputation. This condition is especially relevant for military combat injuries, where the rates of amputations are higher than the general population [19]. In recent United States military conflicts in Iraq and Afghanistan, the incidence rate of amputation is estimated at 5.29 per 100,000 deployed troops [20]. HO occurs after traumatic and combat-related amputations and has been frequently noted as a potential cause of residual limb pain in amputees [19]. HO has proven to be a problematic clinical complication especially for prosthetic wear. In 1 study, HO was present in as many as 63% of all residual limbs postamputation [19]. It was determined that amputation near the injury site as well as a blast mechanism of injury were both significant risk factors for HO development. HO occurring at the site of an amputation can lead to pain in the residual limb as well as other complications, such as poor fitting of the prosthesis. The most effective forms of combating amputation-related HO are prophylaxis with a nonsteroidal anti-inflammatory drug and surgical excision of the bone growth [19]. Less frequent is the appearance of HO in severe burn victims with a varying incidence rate of 0.2% to 5.6% [21]. The association is poorly understood. However, it is suggested that burn injuries involve the recruitment of inflammatory molecules which promote the osteogenic differentiation of local mesenchymal stem cells [21].
HO Associated with Neurogenic Disorders
Severe neurological damage, such as spinal cord injury (SCI), head injuries, strokes, and brain tumors, can result in HO. In SCI, the incidence rate of HO varies from 16% to 53% with all HO occurring below the level of the spinal cord lesion, most commonly at the hip [22]. HO also occurs in 10% to 20% of patients with a closed head injury for whom the ossification mostly develops on the affected side of the brain trauma or stroke [22,23]. Limb spasticity has also been seen to increase the risk of HO development; those patients with extensive neurogenic HO usually present with severe spasticity [22].
Cellular/Molecular Manifestations of HO
HO Progenitors in the Peripheral Nerves
A great deal of controversy has surrounded the actual nature of the osteoprogenitor for HO. It was originally assumed that the progenitor for HO would be the bone marrow mesenchymal stem cell, originally defined by the experiments of Pittenger et al. [24]. These authors showed that cells from bone marrow that preferentially adhered to plastic could be isolated and shown to differentiate to osteoblasts, chondrocytes, and adipocytes. However, early published reports indicated that the progenitors for HO were derived locally rather than from bone marrow [25,26]. Then 3 reports appeared, 1 describing the endothelial origin of osteoprogenitors for HO [27], another noting that the progenitor was similar to those of skeletal muscle [28], and the third noting the neural origin of HO osteoprogenitors [5]. Interestingly, although these studies disagreed on the tissue of origin, they agreed upon the characteristic markers exhibited by the osteoprogenitor, which included Tie2, SP7 (osterix), and PDGFRA. Despite this controversy, it is generally agreed that HO is preceded by a period of neuroinflammation [4,5], where substance P and TRPV1 are expressed. If either of these pathways are blocked (eg, substance P binding to its receptor or TRPV1), then heterotopic bone formation is substantially reduced [4,5].
Recent studies in mice showed that the osteoprogenitors can be identified inthe endoneurium of peripheral nerves [10]. They can exit the nerve via the endoneurial vasculature, cross the blood-nerve barrier, and start to express vascular cell markers Tie2 and claudin 5 (Figure 1). Lineage tracing is a strategy, where a mouse possesses a tamoxifen-inducible Wnt1 (neural) promoter drives Cre recombinase that also possesses a floxed reporter (tomato red). If the animal is given tamoxifen, any cells expressing Wnt1 (neural progenitor marker) will express the red reporter permanently and can then be detected even after expression of Wnt1 has been terminated. In these experiments the osteoblasts were found to possess the red reporter suggesting their neural origin. Chondrocytes and transient brown adipocytes also possessed the reporter indicating that they too had been derived from progenitors within the nerve.
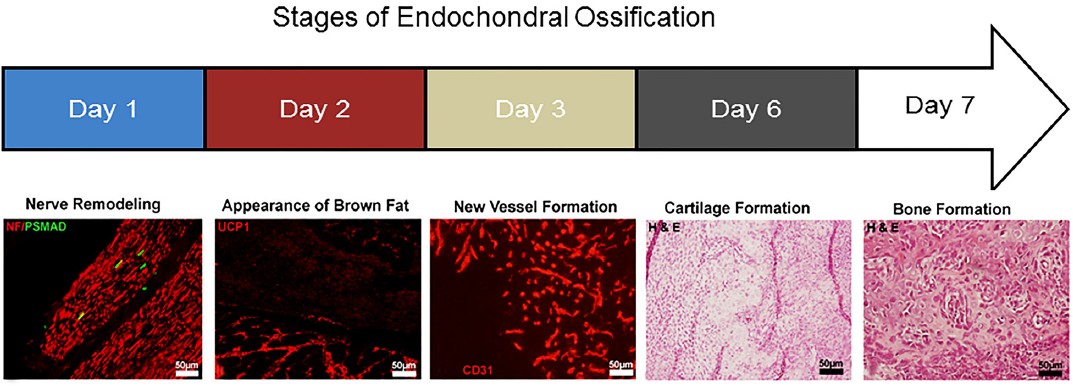
After entering the vasculature, the osteoprogenitors were eventually found to extravasate through the vessel wall to the site of new bone formation. We propose that the expression of these vascular markers is a component of how these cells safely exit the blood-nerve barrier. Recently, it has been shown that traumatic brain injury compromises both the blood-nerve and blood-brain barriers. This may be part of the reason why traumatic brain injury induces HO. Since the cells extravasate from the vessels to reside between muscle fibers, it is easy to see how they could also appear to be localized progentiors potentially derived from the vasculature [28].
We have found that the HO in humans is similar to the murine models that use low levels of BMP2 to induce the bone formation. Analysis of tissues from early human HO shows that many of the cells in the endoneurium or peripheral nerves are positive for phosphoSMADs 1, 5, and 8, indicating that these cells have been activated by BMP. Additionally, osterix is expressed in many of the cells in the endoneurium, and in nerves that are trapped in the newly formed bone. The bone adjacent to these nerves is replete with osteoblasts that bear the neural markers found in the nerves that are directly adjacent. Finally, osteoblasts associated with bone, appear to also express the tight junction molecule claudin 5. In the mouse model we have also recently found that after BMP2 induction, there is axonal sprouting that paves the way for the innervation and vascularization of the newly forming bone. The tBAT [6,7] has several tasks to perform including making the initial microenvironment hypoxic [6] for nerve growth [29] and cartilage formation [6], followed by induction of vascularization [9], which brings the osteoprogenitors to the site of bone formation [10]. Figure 2 shows a schematic representation of the process.
HO and the Blood-Nerve Barrier
Developing a reliable model of HO in larger animals for regenerative medicine has been challenging for investigators. Most HO models involve the delivery or in vivo expression of BMP2 at a variety of concentrations. However, as the size and complexity of the nerve structures increase in species, the process becomes more unreliable, most likely due to the problems associated with getting the BMP2 into the endoneurium of the nerve. The biological differences associated with the complexity of the perineurium and the blood-nerve barrier could explain the differences observed in modeling HO in larger animals. Activated matrix metalloproteinase-9 (MMP-9) is a protease that has the ability to open the blood-nerve barrier. It has been shown to be able to open the blood-nerve barrier in studies of dementia [30] and multiple sclerosis [31]. Studies that have compared the ability of MMP-9 to open the blood-nerve barrier in the central nervous system and the peripheral nervous system have found that while the mechanism and timing is different, MMP-9 induction was observed in both models. Therefore, it is not surprising that when larger animals were found to not express MMP-9, but rather a pro- and uncleaved version of the protease, HO characterized to be separate and distal from the skeletal bone did not occur [32]. MMP-9 was suggested to play a critical role in the opening of the blood-nerve barrier to allow progenitors in the endoneurium to respond to BMP2 expression.
HO and the Blood-Nerve Barrier
Developing a reliable model of HO in larger animals for regenerative medicine has been challenging for investigators. Most HO models involve the delivery or in vivo expression of BMP2 at a variety of concentrations. However, as the size and complexity of the nerve structures increase in species, the process becomes more unreliable, most likely due to the problems associated with getting the BMP2 into the endoneurium of the nerve. The biological differences associated with the complexity of the perineurium and the blood-nerve barrier could explain the differences observed in modeling HO in larger animals. Activated matrix metalloproteinase-9 (MMP-9) is a protease that has the ability to open the blood-nerve barrier. It has been shown to be able to open the blood-nerve barrier in studies of dementia [30] and multiple sclerosis [31]. Studies that have compared the ability of MMP-9 to open the blood-nerve barrier in the central nervous system and the peripheral nervous system have found that while the mechanism and timing is different, MMP-9 induction was observed in both models. Therefore, it is not surprising that when larger animals were found to not express MMP-9, but rather a pro- and uncleaved version of the protease, HO characterized to be separate and distal from the skeletal bone did not occur [32]. MMP-9 was suggested to play a critical role in the opening of the blood-nerve barrier to allow progenitors in the endoneurium to respond to BMP2 expression.
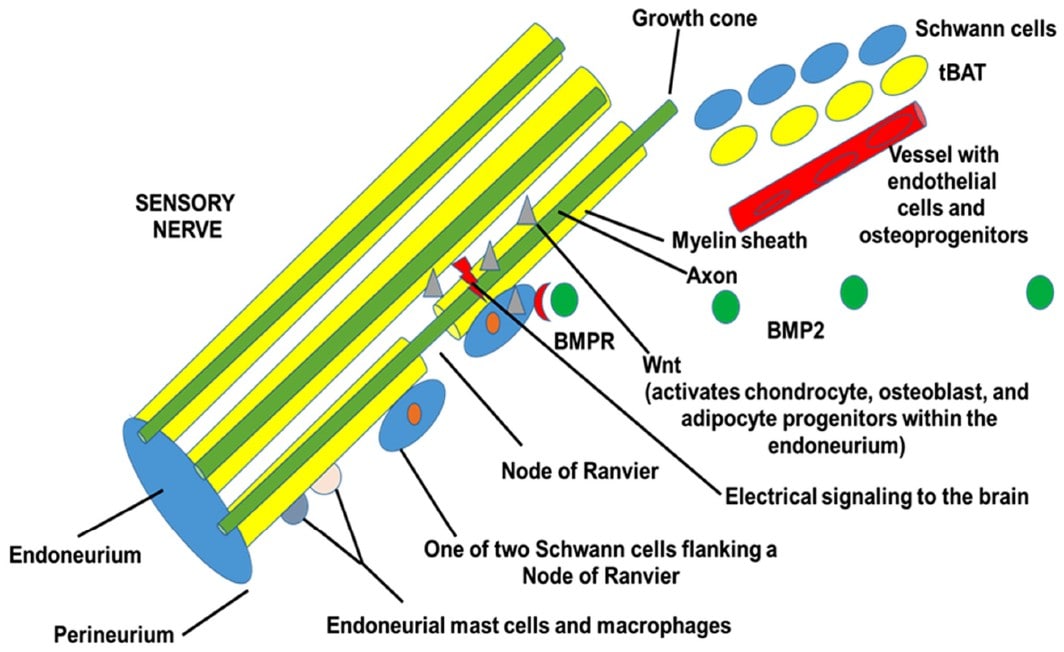
Plasminogen, a protein that binds blood platelets and cleaves MMP-9, has been shown to have effects on bone formation. Plasminogen can only be cleaved to its active form plasmin when platelets are activated and recruited to the tissues. Studies of osteogenesis imperfecta have described the changes in bone formation when platelet-rich plasma is administered to the patient [33]. Finally, studies show that the expression of α2-Antiplasmin leads to bone loss [34]. Collectively, the data suggest that activation and recruitment of blood platelets to the tissues is required for HO.
Consistent with these studies, it has been found that fibrinolysis is needed for HO. When a fibrin matrix was added to plasminogen-deficient mice, robust heterotopic and orthotopic ossification was observed, suggesting that fibrinolysis is essential for HO [35]. Thrombin and fibrin, products of the same pathway that activates and cleaves MMP-9, have been shown to begin an immune response that may be critical for bone formation [36]. In addition, fibrinogen degradation products, or products resulting from fibrinolysis, have been shown to cause mast cell degranulation. Mast cell degranulation has been linked to neurogenic inflammation, which is essential for HO [4,5]. Mast cells most likely play a key role in allowing the barrier to open, and blocking their degranulation resulted in blocking HO formation in a murine model [5]. Studies suggest that inflammation is needed for remodeling of the nerve for release of necessary progenitors for cartilage and bone formation [10]. In addition to mast cells, macrophages also play a key role in the clean-up and remodeling of the nerve and surrounding tissues coordinating with this neurovascular unit to start to support regeneration.
In studies where the model lacked activated MMP-9, it was hypothesized that progenitors could be pulled from the periosteum of the skeletal bone to produce de novo bone formation. However, the resulting bone formation could be described as orthotopic rather than heterotopic bone formation [32]. The studies that suggested that fibrinolysis was essential for HO also suggest that this process will prevent fracture repair in plasminogen-deficient mice. While some of the de novo bone formation presented in these studies appears away from the skeletal bone, most of the bone formation appears to be orthotopic [32], suggesting that plasminogen may play a role in the formation of HO.
Consistent with these studies, it has been found that fibrinolysis is needed for HO. When a fibrin matrix was added to plasminogen-deficient mice, robust heterotopic and orthotopic ossification was observed, suggesting that fibrinolysis is essential for HO [35]. Thrombin and fibrin, products of the same pathway that activates and cleaves MMP-9, have been shown to begin an immune response that may be critical for bone formation [36]. In addition, fibrinogen degradation products, or products resulting from fibrinolysis, have been shown to cause mast cell degranulation. Mast cell degranulation has been linked to neurogenic inflammation, which is essential for HO [4,5]. Mast cells most likely play a key role in allowing the barrier to open, and blocking their degranulation resulted in blocking HO formation in a murine model [5]. Studies suggest that inflammation is needed for remodeling of the nerve for release of necessary progenitors for cartilage and bone formation [10]. In addition to mast cells, macrophages also play a key role in the clean-up and remodeling of the nerve and surrounding tissues coordinating with this neurovascular unit to start to support regeneration.
In studies where the model lacked activated MMP-9, it was hypothesized that progenitors could be pulled from the periosteum of the skeletal bone to produce de novo bone formation. However, the resulting bone formation could be described as orthotopic rather than heterotopic bone formation [32]. The studies that suggested that fibrinolysis was essential for HO also suggest that this process will prevent fracture repair in plasminogen-deficient mice. While some of the de novo bone formation presented in these studies appears away from the skeletal bone, most of the bone formation appears to be orthotopic [32], suggesting that plasminogen may play a role in the formation of HO.
As the molecular and cellular mechanism of HO is further dissected, the role of peripheral nerves starts to become more apparent. With a greater understanding of this mechanism, surgeons can start to have additional information for assessing risk in their patients, and, potentially, leading to improved methods for early intervention to prevent HO without disrupting bone healing and tissue repair.
REFERENCES
[1] Pignolo RJ, Shore EM, Kaplan FS. Fibrodysplasia ossificans progressiva: diagnosis, management, and therapeutic horizons. Pediatr Endocrinol Rev. 2013;10(Suppl 2):437-48.
[2] Kaplan FS, Chakkalakal SA, Shore EM. Fibrodysplasia ossificans progressiva: mechanisms and models of skeletal metamorphosis. Dis Model Mech. 2012;5:756-62.
[3] Cushner FD, Morwessel RM. Myositis ossificans traumatica. Orthop Rev. 1992;21:1319-26.
[4] Kan L, Lounev VY, Pignolo RJ, Duan L, Liu Y, Stock SR, McGuire TL, Lu B, Gerard NP, Shore EM, Kaplan FS, Kessler JA. Substance P signaling mediates BMP-dependent heterotopic ossification. J Cell Biochem. 2011;112:2759-72.
[5] Salisbury E, Rodenberg E, Sonnet C, Hipp J, Gannon FH, Vadakkan TJ, Dickinson ME, Olmsted-Davis EA, Davis AR. Sensory nerve induced inflammation contributes to heterotopic ossification. J Cell Biochem. 2011;112:2748-58.
[6] Olmsted-Davis E, Gannon FH, Ozen M, Ittmann MM, Gugala Z, Hipp JA, Moran KM, Fouletier-Dilling CM, Schumara-Martin S, Lindsey RW, Heggeness MH, Brenner MK, Davis AR.. Hypoxic adipocytes pattern early heterotopic bone formation. Am J Pathol. 2007;170:620-32.
[7] Salisbury EA, Lazard ZW, Ubogu EE, Davis AR, Olmsted-Davis EA. Transient brown adipocyte-like cells derive from peripheral nerve progenitors in response to bone morphogenetic protein 2. Stem Cells Transl Med. 2012;1:874-85.
[8] Ranganathan K, Loder S, Agarwal S, Wong VW, Forsberg J, Davis TA, Wang S, James AW, Levi B. Heterotopic ossification: basic-science principles and clinical correlates. J Bone Joint Surg Am. 2015;97:1101-11.
[9] Dilling CF, Wada AM, Lazard ZW, Salisbury EA, Gannon FH, Vadakkan TJ, Gao L, Hirschi K, Dickinson ME, Davis AR, Olmsted-Davis EA. Vessel formation is induced prior to the appearance of cartilage in BMP-2-mediated heterotopic ossification. J Bone Miner Res. 25:1147-56.
[10] Lazard ZW, Olmsted-Davis EA, Salisbury EA, Gugala Z, Sonnet C, Davis EL, Beal E, Ubogu EE, Davis AR. 2015. Osteoblasts have a neural origin in heterotopic ossification. Clin Orthop Relat Res. 2015;473:2790-806.
[11] Shore EM. Fibrodysplasia ossificans progressiva: a human genetic disorder of extraskeletal bone formation, or how does one tissue become another? Wiley Interdiscip Rev Dev Biol. 2012;1:153-65.
[12] Beiner JM, Jokl P. Muscle contusion injury and myositis ossificans traumatica. Clin Orthop Relat Res. 2002;403:S110-9.
[13] Ootes D, Lambers KT, Ring DC. The epidemiology of upper extremity injuries presenting to the emergency department in the United States. Hand (N Y). 2012;7:18-22.
[14] Hong CC, Nashi N, Hey HW, Chee YH, Murphy D. Clinically relevant heterotopic ossification after elbow fracture surgery: a risk factors study. Orthop Traumatol Surg Res. 2015;101:209-13.
[15] Foruria AM, Marti M, Sanchez-Sotelo J. Proximal humeral fractures treated conservatively settle during fracture healing. J Orthop Trauma. 2015;29:e24-30.
[16] Bauer AS, Lawson BK, Bliss RL, Dyer GS. Risk factors for posttraumatic heterotopic ossification of the elbow: case-control study. J Hand Surg Am. 2012;37:1422-9.
[17] Firoozabadi R, O'Mara TJ, Swenson A, Agel J, Beck JD, Routt M. Risk factors for the development of heterotopic ossification after acetabular fracture fixation. Clin Orthop Relat Res. 2014;472:3383-8.
[18] Kocic M, Lazovic M, Mitkovic M, Djokic B. Clinical significance of the heterotopic ossification after total hip arthroplasty. Orthopedics. 2010;33:16.
[19] Potter BK, Burns TC, Lacap AP, Granville RR, Gajewski DA. Heterotopic ossification following traumatic and combat-related amputations. Prevalence, risk factors, and preliminary results of excision. J Bone Joint Surg Am. 2007;89:476-86.
[20] Krueger CA, Wenke JC, Ficke JR. Ten years at war: comprehensive analysis of amputation trends. J Trauma Acute Care Surg. 2012;73:S438-44.
[21] Schneider JC, Simko LC, Goldstein R, Shie VL, Chernack B, Levi B, Jayakumar P, Kowalske KJ, Herndon DN, Gibran NS, Ryan CM. Predicting heterotopic ossification early after burn injuries: a risk scoring system. Ann Surg. 2016. [Epub ahead of print].
[22] van Kuijk AA, Geurts AC, van Kuppevelt HJ. Neurogenic heterotopic ossification in spinal cord injury. Spinal Cord. 2002;40:313-26.
[23] Mavrogenis AF, Soucacos PN, Papagelopoulos PJ. Heterotopic ossification revisited. Orthopedics. 2011;34:177.
[24] Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S, Marshak DR. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284:143-7.
[25] Hegyi L, Gannon FH, Glaser DL, Shore EM, Kaplan FS, Shanahan CM. Stromal cells of fibrodysplasia ossificans progressiva lesions express smooth muscle lineage markers and the osteogenic transcription factor Runx2/Cbfa-1: clues to a vascular origin of heterotopic ossification? J Pathol. 2003;201:141-8.
[26] Lounev VY, Ramachandran R, Wosczyna MN, Yamamoto M, Maidment AD, Shore EM, Glaser DL, Goldhamer DJ, Kaplan FS. Identification of progenitor cells that contribute to heterotopic skeletogenesis. J Bone Joint Surg Am. 2009;91:652-63.
[27] Medici D, Shore EM, Lounev VY, Kaplan FS, Kalluri R, Olsen BR. Conversion of vascular endothelial cells into multipotent stem-like cells. Nat Med. 2010;16:1400-6.
[28] Wosczyna MN, Biswas AA, Cogswell CA, Goldhamer DJ. Multipotent progenitors resident in the skeletal muscle interstitium exhibit robust BMP-dependent osteogenic activity and mediate heterotopic ossification. J Bone Miner Res. 2012;27:1004-17.
[29] Cattin AL, Burden JJ, Van Emmenis L, Mackenzie FE, Hoving JJ, Garcia Calavia N, Guo Y, McLaughlin M, Rosenberg LH, Quereda V, Jamecna D, Napoli I, Parrinello S1, Enver T, Ruhrberg C, Lloyd AC. Macrophage-induced blood vessels guide Schwann cell-mediated regeneration of peripheral nerves. Cell. 2015;162:1127-39.
[30] Weekman EM, Wilcock DM. Matrix metalloproteinase in blood-brain barrier breakdown in dementia. J Alzheimers Dis. 2015;49:893-903.
[31] Rahimi Z, Abdan Z, Rahimi Z, Razazian N, Shiri H, Vaisi-Raygani A, Shakiba E, Vessal M, Moradi MT. Functional promoter polymorphisms of MMP-2 C-735T and MMP-9 C-1562T and their synergism with MMP-7 A-181G in multiple sclerosis. Immunol Invest. 2016;45:543-52.
[32] Davis EL, Sonnet C, Lazard ZW, Henslee G, Gugala Z, Salisbury EA, Strecker EV, Davis TA, Forsberg JA, Davis AR, Olmsted-Davis EA. Location-dependent heterotopic ossification in the rat model: The role of activated matrix metalloproteinase 9. J Orthop Res. 2016;34(11):1894-904.
[33] Bishop N. Bone material properties in osteogenesis imperfecta. J Bone Miner Res. 2016;31:699-708.
[34] Shiomi A, Kawao N, Yano M, Okada K, Tamura Y, Okumoto K, Matsuo O, Akagi M, Kaji H. Alpha(2)-Antiplasmin is involved in bone loss induced by ovariectomy in mice. Bone. 2015;79:233-41.
[35] Yuasa M, Mignemi NA, Nyman JS, Duvall CL, Schwartz HS, Okawa A, Yoshii T, Bhattacharjee G, Zhao C, Bible JE, Obremskey WT, Flick MJ, Degen JL, Barnett JV, Cates JM, Schoenecker JG. Fibrinolysis is essential for fracture repair and prevention of heterotopic ossification. J Clin Invest. 2015;125:3117-31.
[36] Szaba FM, Smiley ST. Roles for thrombin and fibrin(ogen) in cytokine/chemokine production and macrophage adhesion in vivo. Blood. 2002;99:1053-9.
[1] Pignolo RJ, Shore EM, Kaplan FS. Fibrodysplasia ossificans progressiva: diagnosis, management, and therapeutic horizons. Pediatr Endocrinol Rev. 2013;10(Suppl 2):437-48.
[2] Kaplan FS, Chakkalakal SA, Shore EM. Fibrodysplasia ossificans progressiva: mechanisms and models of skeletal metamorphosis. Dis Model Mech. 2012;5:756-62.
[3] Cushner FD, Morwessel RM. Myositis ossificans traumatica. Orthop Rev. 1992;21:1319-26.
[4] Kan L, Lounev VY, Pignolo RJ, Duan L, Liu Y, Stock SR, McGuire TL, Lu B, Gerard NP, Shore EM, Kaplan FS, Kessler JA. Substance P signaling mediates BMP-dependent heterotopic ossification. J Cell Biochem. 2011;112:2759-72.
[5] Salisbury E, Rodenberg E, Sonnet C, Hipp J, Gannon FH, Vadakkan TJ, Dickinson ME, Olmsted-Davis EA, Davis AR. Sensory nerve induced inflammation contributes to heterotopic ossification. J Cell Biochem. 2011;112:2748-58.
[6] Olmsted-Davis E, Gannon FH, Ozen M, Ittmann MM, Gugala Z, Hipp JA, Moran KM, Fouletier-Dilling CM, Schumara-Martin S, Lindsey RW, Heggeness MH, Brenner MK, Davis AR.. Hypoxic adipocytes pattern early heterotopic bone formation. Am J Pathol. 2007;170:620-32.
[7] Salisbury EA, Lazard ZW, Ubogu EE, Davis AR, Olmsted-Davis EA. Transient brown adipocyte-like cells derive from peripheral nerve progenitors in response to bone morphogenetic protein 2. Stem Cells Transl Med. 2012;1:874-85.
[8] Ranganathan K, Loder S, Agarwal S, Wong VW, Forsberg J, Davis TA, Wang S, James AW, Levi B. Heterotopic ossification: basic-science principles and clinical correlates. J Bone Joint Surg Am. 2015;97:1101-11.
[9] Dilling CF, Wada AM, Lazard ZW, Salisbury EA, Gannon FH, Vadakkan TJ, Gao L, Hirschi K, Dickinson ME, Davis AR, Olmsted-Davis EA. Vessel formation is induced prior to the appearance of cartilage in BMP-2-mediated heterotopic ossification. J Bone Miner Res. 25:1147-56.
[10] Lazard ZW, Olmsted-Davis EA, Salisbury EA, Gugala Z, Sonnet C, Davis EL, Beal E, Ubogu EE, Davis AR. 2015. Osteoblasts have a neural origin in heterotopic ossification. Clin Orthop Relat Res. 2015;473:2790-806.
[11] Shore EM. Fibrodysplasia ossificans progressiva: a human genetic disorder of extraskeletal bone formation, or how does one tissue become another? Wiley Interdiscip Rev Dev Biol. 2012;1:153-65.
[12] Beiner JM, Jokl P. Muscle contusion injury and myositis ossificans traumatica. Clin Orthop Relat Res. 2002;403:S110-9.
[13] Ootes D, Lambers KT, Ring DC. The epidemiology of upper extremity injuries presenting to the emergency department in the United States. Hand (N Y). 2012;7:18-22.
[14] Hong CC, Nashi N, Hey HW, Chee YH, Murphy D. Clinically relevant heterotopic ossification after elbow fracture surgery: a risk factors study. Orthop Traumatol Surg Res. 2015;101:209-13.
[15] Foruria AM, Marti M, Sanchez-Sotelo J. Proximal humeral fractures treated conservatively settle during fracture healing. J Orthop Trauma. 2015;29:e24-30.
[16] Bauer AS, Lawson BK, Bliss RL, Dyer GS. Risk factors for posttraumatic heterotopic ossification of the elbow: case-control study. J Hand Surg Am. 2012;37:1422-9.
[17] Firoozabadi R, O'Mara TJ, Swenson A, Agel J, Beck JD, Routt M. Risk factors for the development of heterotopic ossification after acetabular fracture fixation. Clin Orthop Relat Res. 2014;472:3383-8.
[18] Kocic M, Lazovic M, Mitkovic M, Djokic B. Clinical significance of the heterotopic ossification after total hip arthroplasty. Orthopedics. 2010;33:16.
[19] Potter BK, Burns TC, Lacap AP, Granville RR, Gajewski DA. Heterotopic ossification following traumatic and combat-related amputations. Prevalence, risk factors, and preliminary results of excision. J Bone Joint Surg Am. 2007;89:476-86.
[20] Krueger CA, Wenke JC, Ficke JR. Ten years at war: comprehensive analysis of amputation trends. J Trauma Acute Care Surg. 2012;73:S438-44.
[21] Schneider JC, Simko LC, Goldstein R, Shie VL, Chernack B, Levi B, Jayakumar P, Kowalske KJ, Herndon DN, Gibran NS, Ryan CM. Predicting heterotopic ossification early after burn injuries: a risk scoring system. Ann Surg. 2016. [Epub ahead of print].
[22] van Kuijk AA, Geurts AC, van Kuppevelt HJ. Neurogenic heterotopic ossification in spinal cord injury. Spinal Cord. 2002;40:313-26.
[23] Mavrogenis AF, Soucacos PN, Papagelopoulos PJ. Heterotopic ossification revisited. Orthopedics. 2011;34:177.
[24] Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S, Marshak DR. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284:143-7.
[25] Hegyi L, Gannon FH, Glaser DL, Shore EM, Kaplan FS, Shanahan CM. Stromal cells of fibrodysplasia ossificans progressiva lesions express smooth muscle lineage markers and the osteogenic transcription factor Runx2/Cbfa-1: clues to a vascular origin of heterotopic ossification? J Pathol. 2003;201:141-8.
[26] Lounev VY, Ramachandran R, Wosczyna MN, Yamamoto M, Maidment AD, Shore EM, Glaser DL, Goldhamer DJ, Kaplan FS. Identification of progenitor cells that contribute to heterotopic skeletogenesis. J Bone Joint Surg Am. 2009;91:652-63.
[27] Medici D, Shore EM, Lounev VY, Kaplan FS, Kalluri R, Olsen BR. Conversion of vascular endothelial cells into multipotent stem-like cells. Nat Med. 2010;16:1400-6.
[28] Wosczyna MN, Biswas AA, Cogswell CA, Goldhamer DJ. Multipotent progenitors resident in the skeletal muscle interstitium exhibit robust BMP-dependent osteogenic activity and mediate heterotopic ossification. J Bone Miner Res. 2012;27:1004-17.
[29] Cattin AL, Burden JJ, Van Emmenis L, Mackenzie FE, Hoving JJ, Garcia Calavia N, Guo Y, McLaughlin M, Rosenberg LH, Quereda V, Jamecna D, Napoli I, Parrinello S1, Enver T, Ruhrberg C, Lloyd AC. Macrophage-induced blood vessels guide Schwann cell-mediated regeneration of peripheral nerves. Cell. 2015;162:1127-39.
[30] Weekman EM, Wilcock DM. Matrix metalloproteinase in blood-brain barrier breakdown in dementia. J Alzheimers Dis. 2015;49:893-903.
[31] Rahimi Z, Abdan Z, Rahimi Z, Razazian N, Shiri H, Vaisi-Raygani A, Shakiba E, Vessal M, Moradi MT. Functional promoter polymorphisms of MMP-2 C-735T and MMP-9 C-1562T and their synergism with MMP-7 A-181G in multiple sclerosis. Immunol Invest. 2016;45:543-52.
[32] Davis EL, Sonnet C, Lazard ZW, Henslee G, Gugala Z, Salisbury EA, Strecker EV, Davis TA, Forsberg JA, Davis AR, Olmsted-Davis EA. Location-dependent heterotopic ossification in the rat model: The role of activated matrix metalloproteinase 9. J Orthop Res. 2016;34(11):1894-904.
[33] Bishop N. Bone material properties in osteogenesis imperfecta. J Bone Miner Res. 2016;31:699-708.
[34] Shiomi A, Kawao N, Yano M, Okada K, Tamura Y, Okumoto K, Matsuo O, Akagi M, Kaji H. Alpha(2)-Antiplasmin is involved in bone loss induced by ovariectomy in mice. Bone. 2015;79:233-41.
[35] Yuasa M, Mignemi NA, Nyman JS, Duvall CL, Schwartz HS, Okawa A, Yoshii T, Bhattacharjee G, Zhao C, Bible JE, Obremskey WT, Flick MJ, Degen JL, Barnett JV, Cates JM, Schoenecker JG. Fibrinolysis is essential for fracture repair and prevention of heterotopic ossification. J Clin Invest. 2015;125:3117-31.
[36] Szaba FM, Smiley ST. Roles for thrombin and fibrin(ogen) in cytokine/chemokine production and macrophage adhesion in vivo. Blood. 2002;99:1053-9.